The Evolution of Ageing
Formal models of ageing via damage accumulation
Feb 2024 - Present
Supervisors: Prof. Hanna Kokko
So far, we have built some general models to study how selective disappearance induced by stochastic mortality risk alters the predictions of failure accumulation models. Specifically, we start with a general birth-death framework where the number of failed sub-systems within an individual either increases or decreases stochastically due to failure and repair respectively. Mortality is modelled as an additional stochastic rate at which each individual transitions to an unobserved 'cemetery state' and thus disappears from the population. Using diffusion approximations, we then analytically derive expressions showing how selective disappearance systematically alters observed failure and mortality accumulation curves. A fundamental result from mathematics called the Feynman-Kac formula lets us relate measurements made on organisms how measurements made on populations with selective disappearance can be represented in terms of the failure accumulation process alone where mortality is disallowed. Along the way, we also propose some tentative criteria that failure accumulation and mortality rates should obey based on biological principles and show that these criteria generically lead to Gompertz-Makeham and logistic (Perks) mortality curves during the initial part of the failure accumulation process. This work is accessible as a preprint on bioRxiv (Bhat and Kokko, 2026a).
I am now trying to extend these kinds of models to allow for variable finite cohort/sample sizes, systematic differences in individual 'quality', and births of individuals, bringing in the possibility of studying eco-evolutionary dynamics for such systems by tracking mortality patterns over multiple generations. I am trying to do this via certain measure-valued birth-death processes. Below, I provide an outline of the general flavour of models I am currently thinking about.
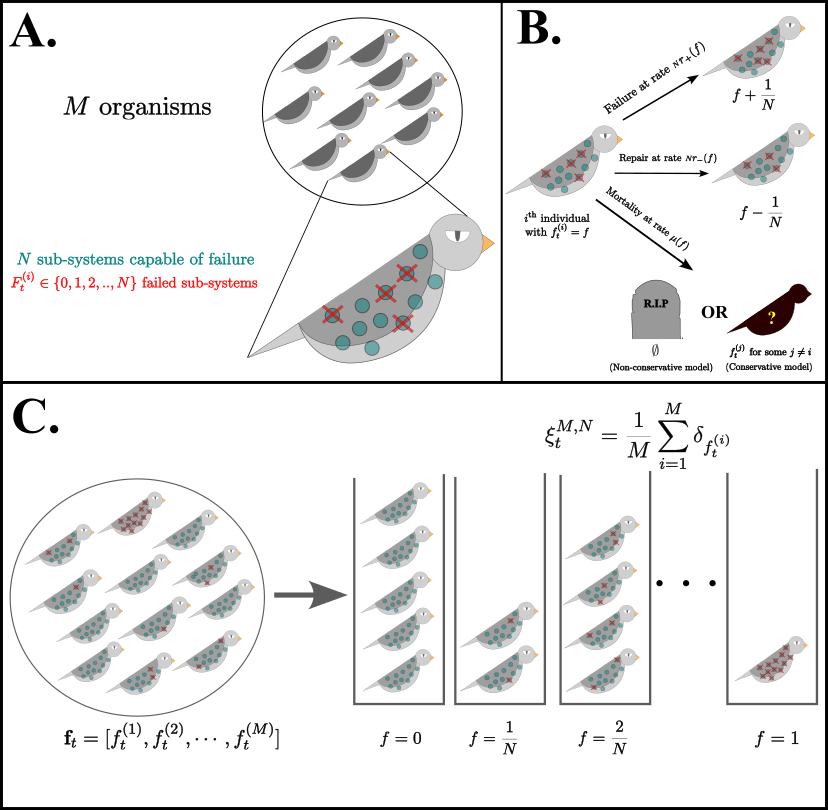
Consider a cohort consisting of \(M\) independent, non-interacting organisms. Each organism has \(N\) sub-systems capable of failure.
Within the \(i\)th organism, \(F^{(i)}_t\), the number of failed sub-systems at time \(t\), follows a density-dependent birth-death process on \(\{0,1,2,\cdots,N\}\) with transition rates \(F \to F \pm 1 \textrm{ at rate } Nr_{\pm}(F/N)\), corresponding to stochastic failure and repair of failed sub-systems. This defines the individual-level dynamics. At the population level, An organism with \(F \in \{0,1,2,\ldots,N\}\) failures is chosen to disappear at rate \(\mu(F/N)\). When an organism disappears, with probability \(\varrho\), it is instantaneously replaced by a copy of another organism chosen uniformly at random from the set of organism that are currently present in the population. We can reframe this as a sub-probability measure-valued stochastic process with state variable
\[\xi^{M,N}_t(\cdot) = \frac{1}{M}\sum\limits_{i=1}^{M}\delta_{f^{(i)}_t}(\cdot)\]where \(f^{(i)}_t = F^{(i)}_t/N\) is a rescaled version of the individual-level process governing failure dynamics. The stochastic process \(\{\xi^{M,N}_t\}_{t \geq 0}\) tracks the proportion of individuals in the population having a proportion \(f = F/N\) failed sub-systems, and takes values in the space of sub-probability measures on \(\{0,\frac{1}{N},\frac{2}{N},\ldots,1\}\).
If we first take \(M \to \infty\) (very large cohort size) and then use a diffusion approximation on \(N\) by neglecting terms of \(\mathcal{O}(N^{-2})\), it can be shown that the stochastic process \(\{\xi^{M,N}_t\}_{t \geq 0}\) converges to a deterministic measure \(P(f,t)\) described by the Fokker-Planck-Kolmogorov type PDE
\[\frac{\partial P(f,t)}{\partial t} = -\frac{\partial}{\partial f}\{r(f)P(f,t)\} + \frac{1}{2N}\frac{\partial^2}{\partial f^2}\{\tau(f)P(f,t)\} - \left[(1-\varrho)\mu(f) +\varrho\left(\mu(f) - \widehat{\mu}(t)\right)\right]P(f,t)\]where \(r(f) = r_{+}(f) - r_-(f), \tau(f) = r_{+}(f) + r_-(f)\), and
\[\widehat{\mu}(t) = \frac{\displaystyle \int\limits_{0}^{1} \mu(x)P(x,t)dx}{\displaystyle \int\limits_{0}^{1} P(x,t)dx}.\]Curiously, from a stochastic process perspective, the PDE above with \(\varrho = 0\) is precisely the Kolmogorov forward equation for the stochastic process (diffusion with killing) we formulated in Bhat and Kokko, 2026a. The PDE with \(\varrho = 1\) describes the conditional probability density of this same stochastic process conditioned on non-absorption/survival. Very similar equations also arise in the multi-level selection literature (see for instance Luo 2014).
In other words, our mathematical formalism lets us uncover a surprising link between the mathematical descriptions of senescence and multi-level selection, despite these phenomena not appearing to have much in common at first glance! To verbally make the connection, observe that defectors spread within a group consisting of cooperators and defectors; when groups compete against each other, defector-rich groups suffer, and between-group selection causes such groups to be systematically under-represented. Exactly analogously, senescing individuals accumulate damage to physiological sub-systems, and ‘damage begets damage’; individuals who are more damaged are more likely to die, hence damage-rich individuals are systematically under-represented in later age classes. Thus, emergent senescence patterns in complex, integrated organisms are formally equivalent to the patterns generated by a within-generation multi-level selection process in which intra-organismal sub-systems play the role of particles, organisms play the role of collectives, and selective disappearance plays the role of group selection. This verbal analogy is made precise in Bhat and Kokko, 2026b.